
Par Serge Poumarat
Depuis trois siècle environ, l'homme a cherché à délimiter et nommer les organismes vivants. Dans le même temps, il essaye de les organiser en un système de classification dit "naturel". La systématique recherche les affinités réelles entre les taxons actuels ou disparus et les regroupe dans un système de classification.
Nomenclature
Pour
nommer une espèce ou un un groupement d'organismes quelconque,
les mycologues doivent suivre des règles contenues dans un
code de nomenclature botanique révisé et édité
tous les quatre ans. Le dernier en date est le code de Vienne
(2006).
Le code international de nomenclature botanique impose
des divisions
officielles telles que "genre", "famille",
"ordres" par exemple.
Systématique
La
systématique essaye de regrouper ente eux les organismes tous
issus d'un même et unique ancêtre dans des taxons
(ensembles d'organismes vivants ou ayant vécu de même
rang systématique). Pour cela elle utilise diverses méthodes
dont la plus puissante à l'heure actuelle est la
"cladistique".
Cette recherche est loin d'être terminée et entraîne
continuellement des bouleversements dans nos connaissances et,
malheureusement, aussi dans les noms des champignons. En effet, les
noms de ceux-ci sont liés au genre, et il suffit que, nos
connaissances s'affinant, certains genres jusque là établis
sur certains critères s'avérant en définitif peu
pertinents, soient reconnus polyphylétiques : toutes les
espèces sensées appartenir à ce genre n'ont pas
toutes le même ancêtre commun. Certaines des espèces
de ce genre vont être "transférées"
dans un ou plusieurs autres genres avec comme conséquences des
changements de noms, certes très gênants mais traduisant
notre plus grande compréhension de la nature. Par exemple, il
a été prouvé récemment que quelques
espèces du genre Macrolepiota (Macrolepiota rachodes, M.
venenata, M. olivieri, M. brunnea...) n'ont pas d'affinités
réelles, non seulement avec le genre Macrolepiota, mais aussi
avec l'ensemble des Lepiotaceae. Leur vraie place est chez les
Agaricaceae dans le genre Chlorophyllum avec l'espèce
Chlorophyllum molybdites. Cela entraîne automatiquement les
changements de nom suivants : Chlorophyllum rhachodes, Chlorophyllum
venenata, etc... Bien que, normalement le genre prioritaire pour
accueillir toutes ces espèces soit le genre sécotoïde
Endoptycum...mais ceci est toute une autre histoire...de
nomenclature!
Les grandes lignes de la classification actuelle
D'après de récentes études (A. A., 2006 ; Hibbet & al., 2007), dont Vizzini (2006) et Durrieru (2008) se sont fait récemment l'écho.
En
premier lieu, certains organismes traditionnellement étudiés
par les mycologues ne font plus partie des champignons (Règne
des Fungi).
Parmi ceux-ci :
-
les Oomycetes,
Hyphochytridiomycetes et Labyrinthulomycetes
font partie du Règne des Chromista
du
Super Règne des Chromoalveolata.
-
les Myxomycètes
font partie du règne des Eumycetozoa
du Super Règne des Amoebozoa.
Règne Fungi (du Super Règne des Opisthokonta du Sous Domaine Unikonta du Domaine Eukarya)
Organismes eucaryotes sans plastes, donc hétérotrophes, ne se nourrissant jamais par phagocytose, pas de phase amiboïde ou pseudoplasmodiale (agrégat d'amibes), à paroi squelettique contenant de la chitine et des bêta-glucanes. Synthèse de la lysine par l'intermédiaire de l'acide alpha amino adipique (A. A. A.).
Il est divisé en 7 phylums :
Microsporidiomycota : environ 800 espèces unicellulaires de parasites endocellulaires de vertébrés et d'invertébrés.
* Mycélium non cloisonné :
Blastocladiomycota : environ 180 espèces avec cellules reproductrices mobiles grâce à un flagelle postérieur, munies d'une structure spéciale appelée "nuclear cap".
Chytridiomycota : environ 700 espèces avec cellules reproductrices mobiles grâce à un flagelle postérieur , sans "nucléar cap".
Zygomycota : environ 900 espèces sans cellules mobiles au cours de la reproduction, saprophytes, parasites ou ectomycorhiziennes (ordre des Endogonales pour ces dernières).
Glomeromycota : environ 160 espèces formants des endomycorhizes avec des plantes ligneuses. Très important pour les forêts tropicales, les Ericacées...
* Mycélium cloisonné :
Ascomycota : Reproduction sexuée par des spores (ascospores) formées de manière endogène dans un sporocyste appelé asque. Elle a généralement lieu dans une fructification spécialisée appelée ascome. La paroi des hyphes est lamellaire avec une couche opaque aux électrons à l'extérieur et une couche épaisse transparente aux électrons à l'intérieur, paroi typique des Ascomycota.
Basidiomycota : Reproduction sexuée par des spores (basidiospores) formées de manière exogène par un sporocyste (ou mieux sporophore) appelé baside qui les porte à l'extrémité d'appendices appelés stérigmates. Cette reproduction a lieu dans une fructification spécialisée appelée basidiome. Présence possible de boucles, de dolipores (sauf exceptions) et la paroi des hyphes est à 2 couches lamellaires opaque aux électrons.
3 sous-phylum :
Taphrinomycotina
Entre autre
l'ordre des Taphrinales : parasites de plantes chez lesquelles ils
provoquent souvent des hyperplasies (foliaires en particuliers) :
Taphrina
deformans
responsable de la cloque du pêcher, "balais de sorcière"
sur les feuillus...
Saccharomycotina (= Hemiascomycetes) : comprennent l'important groupe des levures (par ex. Saccharomycetes)
Pezizomycotina
(= Euascomycetes). Plus de 40 000 espèces.
10
classes : (en gras les classes contenant des espèces formant
des ascomes visibles à l'oeil nu étudiées par
les mycologues amateurs).
- Arthoniomycetes
- Dothideomycetes
- Eurotiomycetes
: contient entre autres les Elaphomyces,
Onygena.
-
Laboulbeniomycetes
- Lecanoromycetes : la plupart des
ascomycetes lichénisés s'inscrivent ici.
-
Leotiomycetes
: Helotiales,
Thelebolales, Rhytismatales...
- Lichinomycetes
- Orbiliomycetes
: Orbiliales.
- Pezizomycetes
: environ 1200 espèces. Ordre des Pezizales. par exemple :
Morchella,
Gyromitra, Helvella, Peziza, Tuber, Terfezia...bref
presque tous les ascomycètes qui nous sont familiers!
-
Sordoriomycetes : la majeure partie des "Pyrénomycètes"
se placent ici. Hypocreales, Diaporthales, Sordoriales,
Xylariales...
Phylum
BASIDIOMYCOTA
Environ
23 000 espèces
3 sous-phylum :
Pucciniomycotina
(= Urediniomycotina, Urediniomycetes, Teliomycetes)
Basidiomycota
primitifs, parasites obligatoires de plantes ou d'insectes. environ
7400 esp. Entre autres les ordres suivants :
-
Uredinales
:
présence d'un sore (rassemblement d'organes reproducteurs).
Parasites connus sous le nom de rouilles, causant de graves maladies
aux plantes (Puccinia,
Gymnosporangium,
etc…). Exemple, Puccinia graminis, la rouille du blé
(hôte intermédiaire : l'épine-vinette).
-Septobasidiales
:
parasites d'insectes (relations complexes). Basidiomes corticoïdes.
Ustilaginomycotina
(= Ustilaginomycetes, Ustomycetes)
Absence de dolipore et
souvent de parenthèsome. Environ 1300 esp. Entre autres les
ordres suivants :
-Ustilaginales
: pas de basidiome mais présence d'un sore. Généralement
parasites de plantes connus sous le nom de charbons (Ustilago)
et de caries (Tilletia).
-Exobasidiales
:
Présence d'un hyménium (basides palissadiques).
Parasites de végétaux provoquant des galles
spectaculaires chez les Ericacée (Rhododendron,
Vaccinium…).
Par ex. genre Exobasidium.
Agaricomycotina
(= Hymenomycetes, Basidiomycetes). Environ 13 500 espèces.
Tous
les basidiomycètes qui intéressent les mycologues
amateurs se placent ici.
3 classes :
Tremellomycetes
par
exemple Tremellales
(plus de 70 espèces) : Tremella,
Tremiscus, Pseudohydnum, Sebacina, Exidia…
Dacryomycetes
par
exemple : Calocera, Dacrymyces, Ditiola...
Agaricomycetes
plus de 13 000 espèces
D'abord quelques ordres primitifs :
Ordre Auriculariales : Auricularia, Exidia...
Ordre Sebacinales : Sebacina...
Ordre Cantharellales (Hydnum, Cantharellus, Craterellus, Clavulina, Sistotrema...)
Ordre
Tulasnellales (Tulasnella)
(mais aussi inclus dans les Cantharellales suivant les études)
Ordre
Trechisporales : Trechispora, Sistotremastrum...
2 sous-classes :
- Sous-classe des Phallomycetidae
Phallales (Phallus, Clathrus...)
Gomphales (Gomphus, Clavariadelphus, Ramaria, Gautiera...)
Hysterangiales (Hysterangium, Phallogaster...)
Geastrales (Geastrum, Spherobolus...)
- Sous-classe des Agaricomycetidae
Agaricales (Agaricus, Amanita, Cortinarius, Entoloma, Tricholoma, Fistulina, Schizophyllum, Lycoperdon, Bovista, Tulostoma, Clavaria, Macrotyphula...)
Boletales (Boletus, Chamonixia, Octavianina, Coniophora, Serpula, Paxillus, Gomphidius, Rhizopogon, Suillus, Astreus, Pisolithus, Scleroderma...)
Atheliales
(Athelia,
Byssocorticium...)
Et
quelques ordres, non classés mais proches des
Agaricomycetidae :
Russulales (Arcangellia, Macowanites, Lactarius, Russula, Bondarzewia, Auriscalpium, Hericium, Lentinellus, Stereum, Scutiger, Peniophora, Aleurodiscus...)
Hymenochaetales
Polyporales (Polyporus, Trametes, Fomitopsis, Phaeolus, Sparassis, Lentinus, Panus...)
Thelephorales (Thelephora, Tomentella, Boletopsis, Phellodon, Sarcodon, Hydnellum, Bankera...)
Corticiales (Corticium, Vuilleminia...)
Gloeophyllales (Gloeophyllum, Neolentinus...)
Bibliographie :
A. A., 2006. A
phylogeny for kingdom Fungi, deep hypha issue. Basidiomycota.
Mycologia, 98 (6) : 917-995.
DURRIEU G., 2008. La classification
des champignons (4). Les Basidiomycètes. La Lettre de la SMF,
11 : 1-3.
HIBBETT D. S. & al., 2007. A higher-level
phylogenetic classification of the Fungi. Mycol. Res., 111 (5) :
509-547.
VIZZINI A., 2004 (paru 2006). Il regno dei funghi :
breve prospetto tassonomico. Boll. Gr. Micol. Bres., ns, 47 (3) :
47-57.
L'exemple
de la classe des Agaricomycetes nous prouve qu'on ne peut pas se fier
à la "bobine" des espèces d'un genre pour en
tirer des conséquences systématiques. Pour s'en
persuader, il suffit de regarder la liste des quelques genres
appartenant à l'ordre des Agaricales, des Boletales ou des
Russulales cités plus haut ou de regarder comment sont
répartis les "clavaires" dans plusieurs ordres
éloignés (Gomphales, Cantharellales, Agaricales). Nous
savons maintenant que les anciens regroupements "Gasteromycetes",
"Aphyllophorales", "Agaricomycetes" pour
pratiques qu'ils étaient, sont totalement artificiels.
Il
est très déroutant pour le "bon sens" de
voir, par exemple, certains genres de champignons à lames
comme les genres "omphaloïdes" Rickenella (Rickenella
fibula),
Loreleia (Loreleia marchantiae, anciennement Omphalina marchantiae)
et Contumyces (Contumyces vesuviana, anciennement Omphalina
vesuviana) n'avoir aucun rapport avec le genre Omphalia, ni même
avec l'ordre des Agaricales mais, par contre, avoisinés, dans
l'ordre des Hymenochaetales, des espèces comme Trichaptum
abietinum ou
Cotylidia pannosa.
Mais
aussi déroutant pour l'esprit que cela soit de prime abord, il
n'est pas difficile de saisir l'explication de ces voisinages ou
éloignements mise en évidence par les méthodes
d'investigation modernes.
Prenons l'exemple des Agaricomycetes. La
silhouette pied-chapeau-lames est un caractère archaïque,
donc apparu tôt. A partir d'un lointain ancêtre commun, à
plusieurs reprises
dans l'évolution, donc de manière
totalement indépendante,
certains Agaricomycetes ont perdu cette silhouette pour adopter un
port, par exemple, résupiné avec ou sans pores (a1 et
a2), ou bien clavarioïdes (b1 et b2). Dans le même temps,
les autres ont gardé une silhouette proche du lointain ancêtre
commun. Un petit schéma pour fixer les idées : 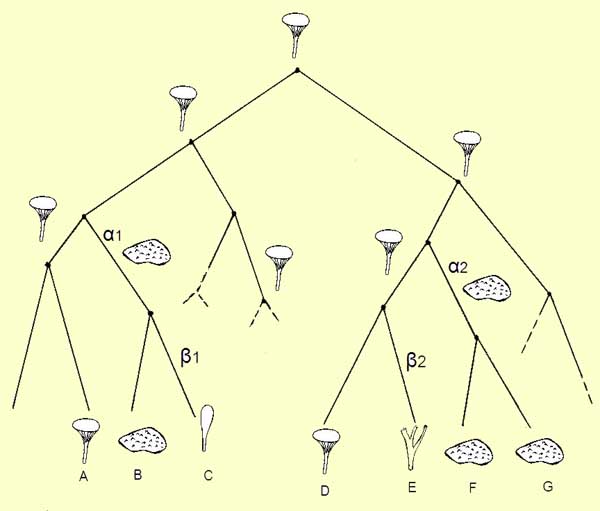
A
l'heure actuelle A et D ont une morphologie très proche, ayant
à peu près concervé celle de leur très
lointain ancêtre commun mais sont éloignés dans
l'évolution au stade actuel. B, F et G d'un côté,
C et E de l'autre, ont un aspect extérieur à peu près
semblable mais par simple convergence
de forme,
étant en réalité tout aussi éloignés
les uns des autres que A et D le sont car il faut "remonter le
temps fort loin pour trouver le premier ancêtre commum. Par
contre, A, B et C d'une part, et D, E, F et G d'autre part, bien que
de formes très dissemblables ont un premier ancêtre
commun moins éloigné dans le temps donc sont
proches.
Un
exemple parlant de convergence de forme pris dans le monde animal :
un dauphin et un requin sont très semblables de forme, ayant
eu à résoudre les mêmes problèmes
hydrodynamiques, mais le premier est en réalité un
mammifère, le second un poisson. Et donc, aussi "déroutant"
que cela puisse paraître à un regard superficiel, le
dauphin est plus proche de l'homme qu'il n'est proche du requin.
De
même, à plusieurs reprises dans l'évolutions, les
Agaricomycetes hypogés se sont enterrés pour diverses
raisons. Toutes les formes sécotioïdes et gastroïdes
ont été regroupées à une époque,
pour leur ressemblance (convergence de forme) dans un ensemble appelé
"Gastéromycèetes". Maintenant, nous savons
que leurs affinités sont, en réalité, très
diverses.
En ce moment même, une espèce à
lames a commencé l'aventure souterraine : Descolea
tenuipes.
Les récoltes semi-épigées à silhouette
classique ont été appelées Descolea
rheophylla
tandis que les récoltes séquestrées, totalement
hypogées, ont été nommées
Setchelliogaster
tenuipes.
La biologie moléculaire a prouvé que c'était
pourtant la même espèce. Ici Descolea tenuipes est
illustrée par la var. rheophylla f. oblongispora (voir NEVILLE
P., POUMARAT S. & IVALDI P., 2005.
Récoltes provençales de Descolea tenuipes (Setch.)
Neville et Poumarat comb. nov. Etude sur la variabilité
sporique. Bull. Soc. Mycol. Fr., 120 (1-4) : 51-71).
- la photo 1
montre des basidiomes étalés ou plus ou moins
comprimés-déformés suivant la profondeur à
laquelle ils poussent dans la litière d'eucalyptus.
- La
photo 2 présente un basidiome normal pour une poussée à
très faible profondeur, dans les tous premiers centimètres
de la litière, et deux basidiomes presque totalement ou
totalement séquestrés qui poussaient à une
profondeur un peu plus grande. Tous ces basidiomes appartiennent
pourtant au même individu.

Descolea tenuipes var. rheophylla f. oblongispora (photo S. Poumarat)
{kind=link}